Об интеллекте, стр. 37
И наконец, есть еще один непрямой способ связи зон коры головного мозга.
Но прежде я хотел бы вам снова напомнить об автоассоциативном свойстве системы памяти (о котором шла речь в главе 2). Как вы, вероятно, помните, автоассоциативные запоминания служат для сохранения последовательностей сигналов. Когда выходной сигнал группы искусственных нейронов передается назад с целью формирования входного потока для всех нейронов (и при этом обратная связь дополняется задержкой), то таким образом сигналы учатся последовательно двигаться друг за другом. По моему мнению, кора головного мозга использует подобный механизм для сохранения последовательностей, но в нем присутствуют еще некоторые особенности. Вместо того чтобы формировать автоассоциативные последовательности из искусственных нейронов, она формирует их из колонок коры головного мозга. Исходящие данные из всех колонок передаются назад, в слой 1. Выходит, слой 1 располагает свежей информацией о том, какие колонки зоны коры головного мозга только что были активными.
Рассмотрим элементы, изображенные на рис. 6.9. Ученым давно известно, что нейроны особо крупных размеров слоя 5 моторной коры (зона M1) непосредственным образом связаны с мышцами и моторными зонами спинного мозга – они обеспечивают управление вашими мышцами и передвижение. Каждый раз, когда вы говорите, печатаете, выполняете любые самые сложные действия, эти клетки посылают высококоординированные импульсы, заставляя сокращаться ваши мышцы.
Совсем недавно ученые открыли, что крупные нейроны слоя 5, возможно, выполняют какие-то функции и в других частях коры головного мозга, а не только в моторных зонах. Например, крупные нейроны слоя 5 зрительной зоны имеют проекции в зоне, отвечающей за движения глаз. Сенсорные зрительные зоны V2 и V4 не только занимаются обработкой зрительных входных сигналов, но также помогают предопределять движение глаз, а следовательно, управляют тем, что вы видите. Крупные нейроны слоя 5 присутствуют в каждой зоне коры головного мозга, что наводит на мысль об их немаловажной роли во всех типах движений.
Аксоны этих нейронов не только участвуют в формировании поведения, они разветвляются надвое. Одно ответвление идет к таламусу, изображенному на рис. 6.9 в виде овальной фигуры. Таламус человека находится в центре головного мозга, под корой, сверху «старого мозга», и окружен белым веществом. Размером и формой он похож на два маленьких птичьих яйца. Все зоны коры головного мозга направляют многочисленные аксоны в таламус, от него тоже исходят аксоны в обратном направлении. Науке известно достаточно об этих связях, но сам таламус очень сложен по своей структуре, и его роль до сих пор мало изучена. Таламус необходим для того, чтобы вести нормальный образ жизни: его повреждение приводит к устойчивому «растительному» состоянию.
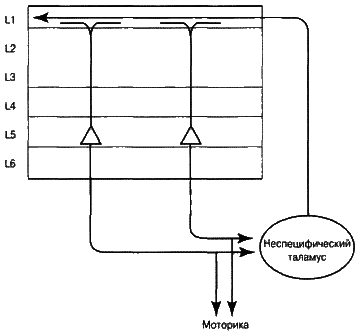
Рис. 6.9. Как посредством таламуса сообщаются текущее состояние и текущее моторное поведение
Существуют несколько путей от таламуса к коре головного мозга, но сейчас лишь один из них представляет для нас интерес. Он начинается с крупных нейронов слоя 5, проецирующихся на группу клеток таламуса, считающихся функционально неспецифическими. Аксоны этих неспецифических клеток идут назад, в слой 1 множества различных зон коры головного мозга. Например, нервные клетки слоя 5 зон V2 и V4 посылают аксоны к таламусу, а таламус, в свою очередь, отправляет информацию назад к слою 1 зон V2 и V4. Аналогичный процесс происходит и в других зонах коры головного мозга. Клетки слоя 5 множества корковых зон посылают сигналы в таламус, который, в свою очередь, отсылает назад информацию в слой 1 этих же самых и связанных с ними зон. Я считаю, что такой круговорот полностью дублирует обратную связь с задержкой, которая позволяет автоассоциативной памяти усваивать последовательности сигналов.
Только что я упомянул о двух источниках входной информации, поступающей в слой 1. Высшие зоны коры головного мозга возбуждают активность в слое 1 более низких зон коры. Активные колонки в пределах одной зоны возбуждают активность в пределах слоя 1 той же зоны через таламус. Эти входящие в слой 1 сигналы можно представить как название песни – считать словами песни (входящие сверху сигналы) и знание, в каком месте песни мы сейчас находимся (задержка в деятельности активных колонок в пределах одной зоны). Таким образом, слой 1 содержит большое количество информации, необходимой нам для прогнозирования времени активизации колонки: название последовательности и наше местонахождение в ней. Используя эти два сигнала слоя 1, зона коры головного мозга в состоянии обучаться и запоминать огромное количество последовательностей сигналов.
Как работает корковая зона: подробности
Не забывая о наших трех схемах (входные сигналы, поступающие вверх по иерархии; разветвляющиеся сигналы, спускающиеся вниз по иерархии; обратная связь с задержкой в таламусе), приступим к рассмотрению того, как зона коры головного мозга выполняет свои функции. Нам необходимо выяснить следующее.
Как зона мозга классифицирует входящие сигналы (вспомните сортировку цветных листов бумаги по ведрам).
Как она усваивает последовательности сигналов (вспомните последовательности интервалов в песне и восприятие лиц посредством комбинаций типа «глаз-нос-глаз»).
Как корковая зона формирует постоянные паттерны, или «имена» последовательностей.
Как она составляет специфические прогнозы (встреча поезда в определенное время или прогнозирование следующего звука мелодии).
Допустим, что колонки одной зоны – это те же ведра, которые мы использовали для воображаемой сортировки получаемых цветных листов бумаги. Каждая колонка помечена точно так же, как и ведро. К клеткам слоя 4 в каждой колонке протянуты волокна ввода из нескольких зон, расположенных ниже по иерархии. Всякий раз, получая правильную комбинацию входных сигналов, они выдают импульсный разряд. Возбуждаясь, клетка слоя 4 «голосует» за то, что полученные сигналы соответствуют своему ярлыку. Точно так же, как при сортировке листов бумаги, входные сигналы могут быть неоднозначными (например несколько колонок могут подходить для единственного входного сигнала). Мы хотим, чтобы зона коры нашего головного мозга сделала выбор: лист бумаги может считаться либо красным, либо оранжевым, но ни в коем случае не тем и другим сразу. Колонка, которой входной сигнал соответствует более всего, должна предотвратить активизацию остальных колонок.
Этим занимаются так называемые тормозные нейроны. Они предотвращают активизацию соседних нервных клеток, эффективно заботясь о том, чтобы был только один «победитель». Эти тормозные нейроны влияют исключительно на зону вокруг «своей» колонки. Другими словами, несмотря на наличие механизма подавления, существует возможность одновременной активизации нескольких колонок. (В мозге ничто не бывает представлено одним единственным нейроном или единственной колонкой.) Для простоты понимания можем представить, что зона выбирает одну и только одну колонку-«победителя», однако на будущее запомним, что на самом деле одновременно активизируются множество колонок. Процесс, используемый корой головного мозга для классификации входных сигналов, сложен и плохо изучен. Я не собираюсь заводить вас в дебри. Вместо этого я предлагаю принять предположение, что зона коры головного мозга отсортировала входной сигнал как активизацию в наборе колонок. Тогда мы сможем сосредоточиться на формировании последовательностей и присвоении им названий.
Каким образом зона коры головного мозга сохраняет последовательность отсортированных сигналов? Я уже излагал свои соображения по этому поводу и сейчас хотел бы углубиться в подробности. Представьте себе, что вы – колонка нервных клеток, а сигнал, поступивший из зоны низшего порядка, заставил активизироваться одну из ваших клеток слоя 4. Вслед за этим возбуждаются клетки в слоях 2 и 3, потом – в слое 5, а затем – в 6-м. Постепенно активизируется вся колонка нейронов. У каждой из клеток слоев 2, 3 и 5 есть тысячи синапсов в слое 1. Если какие-то из этих синапсов активны во время разряда нервных клеток слоев 2, 3 и 5, они усиливаются. Если такое происходит часто, то синапсы слоя 1 становятся достаточно сильными для того, чтобы вызвать вспышки клеток в слоях 2, 3 и 5 даже тогда, когда клетки слоя 4 молчат. Таким образом, некоторые части колонки могут активизироваться, даже если они не получили входного сигнала от более низкой по иерархии зоны коры головного мозга. Именно так клетки слоев 2, 3 и 5 учатся предвосхищать собственную активизацию на основе сигнала, поступающего из слоя 1. «До обучения» колонка может активизироваться исключительно через клетки слоя 4. «После обучения» колонка может частично активизироваться через память. Когда колонка активизируется через синапсы слоя 1, то входящие в нее нейроны предвидят, что произойдет активизация снизу, – это и есть прогноз. Если бы колонка могла разговаривать, она бы сказала: «Когда я активизировалась в прошлом, то как раз этот набор моих синапсов слоя 1 был активен. Как только я увижу такой набор синапсов, сразу же начну генерировать импульс в предвкушении сигнала».