Двойная спираль, стр. 26
Обычно только поздно вечером, вернувшись домой, я пытался разгадать тайну оснований. Их формулы приведены в небольшой книге Дж.Н. Дэвидсон, «Биохимия нуклеиновых кислот», и у меня в Клэр был ее экземпляр. Поэтому я не сомневался, что правильно рисую крохотное изображение оснований. Мне хотелось расположить основания в центре молекулы таким образом, чтобы внешние цепи оказались совершенно регулярными, то есть чтобы сахаро-фосфатные группы каждого нуклеотида имели одинаковую пространственную конфигурацию. Но всякий раз, пытаясь решить эту задачу, я наталкивался на препятствие, заключавшееся в том, что у всех четырех оснований совершенно разная форма. Кроме того, у нас были причины считать, что последовательность оснований в любой полинуклеотидной цепи весьма нерегулярна.
И если просто наугад скручивать две такие цепи, получалась чепуха. Основания покрупнее кое-где должны были соприкасаться, а там, где друг против друга располагались основания поменьше, между ними приходилось оставлять промежуток, ибо соответствующие участки остова недопустимо прогибались. Чтобы этого избежать, нужно было придумать какой-нибудь хитрый прием.
Приходилось ломать голову и над тем, каким образом переплетенные цепи удерживаются вместе благодаря водородным связям между основаниями. Хотя более года назад мы с Фрэнсисом решили, что между основаниями не могут образовываться регулярные водородные связи, теперь стало ясно, что мы были неправы. Наблюдения, показывавшие, что один или несколько атомов водорода в каждом из оснований могут занимать разное положение (таутомерные превращения), заставили нас было прийти к выводу, что все возможные таутомерные формы данного основания встречаются одинаково часто.
Но недавно, перечитывая статьи Дж.М. Гэлланда и Д.О. Джордана о кислотном и щелочном титровании ДНК, я в конце концов убедился в справедливости их заключения, что большинство оснований (если не все) образует водородные связи с другими основаниями. И что еще важнее – эти водородные связи возникают и при очень низких концентрациях ДНК, а значит, они связывают между собой основания, принадлежащие одной и той же молекуле. Кроме того, рентгеновские данные свидетельствовали о том, что все исследованные в чистом виде основания образуют столько нерегулярных водородных связей, сколько допускают законы стереохимии. Таким образом, все дело заключалось в существовании некоего правила, которое управляет образованием водородных связей между основаниями.
На первых порах моя возня с основаниями на бумаге не приносила никаких результатов независимо от того, ходил ли я в тот вечер в кино или нет. Я очень старался очиститься от воспоминаний об «Экстазе», но даже это не помогло обнаружить приемлемые водородные связи, и я заснул, надеясь, что на завтрашней студенческой вечеринке в Даунинге будет много хорошеньких девушек. Но меня ожидало горькое разочарование: там не было никого, кроме пышущих здоровьем спортсменок да двух-трех худосочных светских девиц. Бертран тоже мгновенно понял, что это не для него, и те несколько минут, которые мы из вежливости пробыли там, прежде чем улизнуть, я рассказывал ему, как я пытаюсь обойти отца Питера в состязании за Нобелевскую премию.
Однако нетривиальная идея осенила меня только в середине следующей недели. Она пришла мне в голову, когда я рисовал конденсированные циклы аденина. Я вдруг заметил, какие многообещающие возможности таятся в такой структуре ДНК, где остаток аденина образовывал бы водородные связи, подобные обнаруженным в кристаллах свободного аденина. При этом каждый адениновый остаток соединялся бы двумя водородными связями с таким же остатком, повернутым по отношению к нему на 180°. А главное – подобные симметричные связи могли соединять между собой также и пары гуанинов, цитозинов и тиминов. Я подумал: а что если каждая молекула ДНК состоит из двух цепей с одинаковой последовательностью оснований, а скрепляют эти цепи водородные связи между парами одинаковых оснований? Правда, дело осложнялось тем, что такая структура не может иметь регулярного остова, поскольку пуриновые основания (аденин и гуанин) и пиримидиновые основания (тимин и цитозин) разной формы. На получавшемся остове должны были образовываться небольшие вздутия и впадины в зависимости от того, из пуринов или из пиримидинов состоит центральная пара оснований.
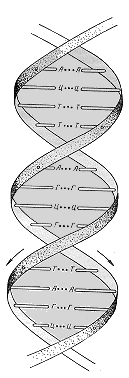
Схематическое изображение молекулы ДНК, построенное с учетом спаривания одинаковых оснований.
Несмотря на то что остов получался такой неаккуратный, у меня забилось сердце. Если ДНК такова, то мое сообщение об этом открытии произведет впечатление разорвавшейся бомбы.
Существование двух переплетенных цепей с одинаковой последовательностью оснований не может быть случайным. Наоборот, это дает право полагать, что одна из цепей каждой молекулы на какой-то более ранней стадии служила матрицей для синтеза другой цепи. По такой схеме репликация гена начиналась бы с его разделения на две одинаковые цепи. Потом на обеих матрицах-родительницах образовывались бы две дочерние цепи, и получались бы две молекулы ДНК, идентичные первоначальной. Таким образом, если бы каждое основание синтезируемой цепи обязательно присоединялось двумя водородными связями к такому же основанию, то это могло бы определять весь ход репликации гена. Правда, в тот вечер я так и не понял, почему к аденину не может присоединиться водородными связями и обычная таутомерная форма гуанина. Могли происходить и другие ошибки в спаривании оснований. Но это меня не очень пугало, поскольку не исключалось участие еще и специфических ферментов. Например, мог существовать такой фермент, который бы заставлял аденин всегда занимать место против остатка аденина на матрице. Был уже первый час ночи, а я увлекался все больше и больше. Нас с Фрэнсисом давно тревожила мысль, что структура ДНК может оказаться внешне очень скучной, ничего не говорящей ни о ее репликации, ни о ее роли в управлении биохимией клетки. И вот теперь, к моему восторгу и изумлению, решение обещало быть чрезвычайно интересным. Больше двух часов я не мог заснуть от радости, и перед моими закрытыми глазами кружились пары адениновых остатков. И только временами меня охватывал страх, что вдруг эта прекрасная идея все-таки неверна.
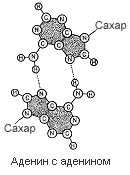
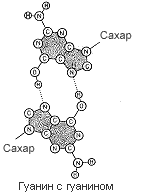
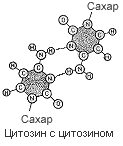
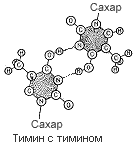
Четыре пары оснований, использованные для постройки модели, в основе которой лежала идея соединения подобного с подобным (пунктиром показаны водородные связи).
26
К полудню следующего дня от моей схемы не осталось камня на камне. Против меня был тот неприятный химический факт, что я выбрал не те таутомерные формы гуанина и тимина. Еще до того, как обнаружилась эта печальная истина, я наспех позавтракал в «Лакомке» и ненадолго вернулся к себе в Клэр, чтобы ответить на письмо Макса Дельбрюка, который сообщал, что специалистам из Калифорнийского технологического института моя статья по генетике бактерий показалась недостаточно обоснованной. Тем не менее, если я этого хочу, он пошлет рукопись в «Труды Национальной Академии наук». Таким образом, я опубликую нелепую идею еще молодым, и у меня будет время одуматься, прежде чем я окончательно утвержусь на гибельном пути.
Сначала это послание возымело желаемое обескураживающее действие. Но теперь, воодушевленный тем, что я, возможно, уже получил самовоспроизводящуюся структуру, я подтвердил, что не сомневаюсь в правильности своих представлений о половом размножении бактерий. Не удержавшись, я добавил, что как раз сегодня изобрел для ДНК изящную структуру, совершенно непохожую на структуру Полинга. Несколько секунд я колебался, не сообщить ли подробности, но решил, что времени на это у меня нет, быстро бросил письмо в почтовый ящик и помчался в лабораторию.